BBC Science Focus, tim peneliti asal Inggris, Afrika Selatan, dan Belanda mengungkapkan bahwa tumbuhan mampu menyerap sekitar 13,12 gigaton karbon dioksida dari atmosfer ke dalam jamur mikoriza setiap tahunnya. Karbon yang diserap ke dalam tanah setara dengan 36% emisi bahan bakar fosil global tahunan. Dunia Sawit pasti sudah mengenal siapa sih Mikoriza itu? Apalagi si jamur ini heboh setelah beberapa lahan sawit terkena serangan Ganoderma. Tak kenal maka tak sayang, yuk kita mengenal lebih jauh tentang Mikoriza ini. Nama “mikoriza” adalah serapan dari istilah bahasa Inggris, mycorrhiza, yang juga bentukan dari dua kata bahasa Yunani Kuno: μύκης mýkēs, “jamur”, dan ῥίζα rhiza.
Selain bermanfaat sebagai mikoriza penyubur tanaman, jamur kapang juga digunakan sebagai ragi pengolah makanan seperti pembuatan roti, tempe, oncom, minuman anggur brewery, dan tapai serta pengobatan antibiotik. sedangkan Kelompok yang menyebabkan keracunan jamur adalah mikotoksin.
Mikoriza adalah cendawan (fungi) yang bersimbiosis dengan tumbuhan. Biasanya simbiosis ini terletak di sistem perakaran tumbuhan. Ada juga cendawan yang bersimbiosis dengan cendawan lainnya, tetapi sebutan mikoriza biasanya ditunjukan untuk cendawan yang melakukan simbiosis dengan tumbuhan. Bentuk simbiosis ini terutama adalah simbiosis mutualisme, meskipun pada beberapa kasus dapat berupa simbiosis parasitisme lemah.
Mikoriza memerlukan akar tumbuhan untuk melengkapi daur hidupnya. Sebaliknya, beberapa tumbuhan bahkan ada yang tergantung pertumbuhannya dengan mikoriza. Beberapa jenis tumbuhan tidak tumbuh atau terhambat pertumbuhannya tanpa kehadiran mikoriza di akarnya. Sebagai misalnya, semaian pinus biasanya gagal tumbuh setelah pemindahan apabila tidak terbentuk jaringan mikoriza di sekitar akarnya.
Mikoriza dapat diinokulasi secara buatan. Namun, inokulasi mikoriza asing memerlukan bantuan mikoriza lokal, misalnya dengan menambahkan tanah dari tempat asal tumbuhan.
Dua kelompok mikoriza terbesar adalah ektomikoriza (EcM) dan endomikoriza (EM). Endomikoriza terutama didominasi oleh mikoriza arbuskular (arbuscular mycorrhizae, AM), ditambah dengan sekelompok mikoriza erikoid dan mikoriza arbutoid yang menginfeksi tumbuhan kelompok Ericoidae.
Semua endomikoriza termasuk ke dalam filum Glomeromycota, misalnya genus Gigaspora, Scultellospora, Acaulospora, Entrophospora, Glomus, dan Sclerocystis. Terdapat sekitar 150 jenis (spesies) spora cendawan AM yang telah dideskripsi. AM tergolong dalam kelompok khusus dari populasi mikoriza yang sangat banyak mengkolonisasi rizosfer, yaitu di dalam akar, permukaan akar, dan di daerah sekitar akar. Hifa eksternal yang berhubungan dengan tanah dan struktur infeksi seperti arbuskula di dalam akar menjamin adanya perluasan penyerapan unsur-unsur hara dari tanah dan peningkatan transfer hara (khususnya fosfor) ke tumbuhan, sedangkan cendawan memperoleh karbon organik dari tumbuhan inangnya (Marschner, 1995).
Mikoriza dibagi menjadi dua kelompok berdasarkan cara menginfeksinya, yaitu ektomikoriza dan endomikoriza. Ektomikoriza menginfeksi permukaan luar tanaman dan di antara sel-sel ujung akar. Akibat serangannya, terlihat jalinan miselia berwarna putih pada bagian rambut-rambut akar, dikenal sebagai jala Hartig. Serangan ini dapat menyebabkan perubahan morfologi akar. Akar-akar memendek, membengkak, bercabang dikotom, dan dapat membentuk pigmen. Infektivitas tergantung isolat dan kultivar tumbuhan inang. Tumbuhan inangnya biasanya tumbuhan tahunan atau pohon. Beberapa di antaranya merupakan komoditas kehutanan dan pertanian seperti sengon, jati, serta beberapa tanaman buah seperti mangga, rambutan, dan jeruk. Selain itu pohon-pohon anggota Betulaceae, Fagaceae, dan Pinaceae juga menjadi inangnya. Pada umumnya ektomikoriza termasuk dalam filum Basidiomycota dan Ascomycota. Ada sedikit anggota Zygomycota yang juga menjadi cendawan ektomikoriza.
Endomikoriza menginfeksi bagian dalam akar, di dalam dan di antara sel-sel ujung akar. Hifa masuk ke dalam sel atau mengisi ruang-ruang antarsel. Jenis mikoriza ini banyak ditemukan pada tumbuhan semusim yang merupakan komoditas pertanian penting, seperti kacang-kacangan, padi, jagung, beberapa jenis sayuran dan tanaman hias. Infeksi ini tidak menyebabkan perubahan morfologi akar, tetapi mengubah penampilan sel dan jaringan akar. Berdasarkan tipe infeksinya, dikenal tiga kelompok endomikoriza: ericaceous (Ericales dengan sejumlah Ascomycota), orchidaceous (Orchidaceae dengan sekelompok Basidiomycota), dan vesikular arbuskular (sejumlah tumbuhan berpembuluh dengan Endogonales, membentuk struktur vesikula (gelembung) dan arbuskula dalam korteks akar) disingkat MVA.
Mikoriza arbuskular (AM, dulu disebut mikoriza vesikular-arbuskular, VAM) tumbuh dari luar perakaran lalu masuk ke dalam jaringan perakaran dan pada gilirannya memasuki sel-sel perakaran. AM di dalam jaringan akan membentuk arbuskula, yaitu jaringan hifa yang menembus sela-sela sel dan bahkan menembus sel melalui plasmalema. Di dalam sel, hifa akan membentuk vesikula, suatu gelembung-gelembung kecil di sitoplasma. AM sulit ditumbuhkan secara aksenik (media buatan) sehingga MVA dianggap merupakan simbion obligat (wajib).
Vesikula berbentuk butiran-butiran di dalam sitoplasma yang mengandung lipid dan menjadi alat reproduksi vegetatif mikoriza, khususnya bila sel pecah akibat rusaknya korteks akar. Arbuskula berwujud kumpulan hifa yang menembus plasmalema dan membantu transportasi hara di dalam sel tumbuhan. Pembentukan vesikula dan arbuskula dalam sel menunjukkan bahwa simbiosis telah terjadi dengan sempurna dan tanaman sudah dapat menikmati hasil kerja sama dengan mikoriza berupa meningkatnya ketersediaan unsur hara yang diserap dari dalam tanah.
Selain vesikula dan arbuskula, terbentuk hifa eksternal yang dapat membantu memperluas ruang penyerapan hara oleh akar. Pada bawang merah, misalnya, panjang hifa eksternal dapat mencapai 80 cm per satu cm panjang akar. Di luar akar, hifa dapat membentuk sporangium yang menghasilkan spora sebagai alat reproduksi.
AM banyak membawa keuntungan bagi tumbuhan simbionnya. Ia memperbaiki hasil tumbuhan dan mengurangi masukan pupuk pada tanaman pertanian. Ini terjadi karena MVA meningkatkan ketersediaan beberapa hara di tanah yang diperlukan tanaman, terutama fosfat. Peningkatan penyerapan fosfat diiringi dengan peningkatan penyerapan hara lain, seperti nitrogen (N), seng (Zn), tembaga (Cu), dan belerang (S).
Selain itu, MVA memperluas ruang tanah yang dapat dijangkau oleh tanaman inang. Jeruk, umpamanya, dikenal responsif terhadap inokulasi MVA. Inokulasi ini dapat mengarah pada menurunnya penggunaan pupuk P. Selain meningkatkan ketersediaan hara, AM meningkatkan toleransi tumbuhan terhadap kurangnya pasokan air. Luasnya jaringan hifa di tanah membantu akar menyerap air. MVA memengaruhi ketahanan tumbuhan inang terhadap serangan penyakit. AM, tergantung jenisnya, dapat mengurangi pengaruh serangan jamur patogen. Demikian pula, juga dapat mengurangi serangan nematoda. Sebaliknya, tumbuhan yang terinfeksi AM menurun ketahanannya terhadap serangan virus.
Pengaruh AM lain yang pernah teramati adalah dukungannya terhadap simbiosis antara bakteri bintil akar dan polong-polongan, produksi giberelin oleh Gibberella mosseae, memengaruhi sintesis fitohormon tertentu, dan memperbaiki struktur agregasi tanah.
Manfaat Umum MVA
Manfaat dari MVA dapat dikelompokkan menjadi tiga, yaitu manfaat dalam ekosistem, manfaat bagi tanaman, dan manfaatnya bagi manusia. Manfaat mikoriza MVA dalam ekosistem sangat penting, yaitu berperan dalam siklus hara, memperbaiki struktur tanah dan menyalurkan karbohidrat dari akar tanaman ke organisme tanah yang lain, sedangkan manfaat bagi tanaman yaitu dapat meningkatkan penyerapan unsur hara, terutama P. MVA ini dapat mengeluarkan enzim fosfatase dan asam-asam organik, khususnya oksalat yang dapat membantu membebaskan P. MVA dapat membantu mengatasi masalah ketersediaan fosfat melalui dua cara, pengaruh langsung melalui jalinan hifa eksternal yang diproduksinya secara intensif sehingga tanaman bermikoriza akan mampu meningkatkan kapasitasnya dalam menyerap unsur hara dan air dan pengaruh tidak langsung, dimana mikoriza dapat memodifikasi fisiologis akar sehingga dapat mengeksresikan asam-asam organik dan fosfatase asam ke dalam tanah. Fosfatase asam merupakan suatu enzim yang dapat mamacu proises mineralisasi P Organik dengan mengkatalisis pelepasan P dari kompleks organik menjadi kompleks anorganik. Manfaat lain yaitu bagi manusia, mikoriza dapat meningkatkan produktivitas tumbuhan, karena dapat memproduksi bunga lebih awal.
Mekanisme Penyerapan Fosfat oleh Mikoriza
Peranan MVA tersebut dalam meningkatkan ketersediaan dan serapan P dan unsur hara lainnya melalui proses sebagai berikut:
Modifikasi Kimia oleh mikoriza dalam proses kelarutan P tanah Pengaruh Mikoriza Arbuskula Pada Ketersediaan dan Penyerapan Unsur Hara Pada tahap ini, terjadi modifikasi kimia oleh mikoriza terhadap akar tanaman, sehingga tanaman mengeksudasi asam-asam norganik dan enzim fosfatase asam yang memacu proses mineralisasi P. Eksudasi akar tersebut terjadi sebagai respon tanaman terhadap kondisi tanah yang kahat P, yang mempengaruhi kimia rizosfer.
Perpendekan jarak difusi oleh tanaman bermikoriza. Mekanisme utama bagi pergerakan P ke permukaan akarah melalui difusi yang terjadi akibat adanya gradien konsentrasi, serta merupakan proses yang sangat lambat. Jarak difusi ion-ion fosfat tersebut dapat diperpendek dengan hifa eksternal CMA, yang juga dapat berfungsi sebagai alat penyerap dan translokasi fosfat.
Penyerapan P tetap terjadi pada tanaman bermikoriza meskipun terjadi penurunan konsentrasi minimum P. Konsentrasi P yang ada di larutan tanah dapat menjadi sangat rendah dan mencapai konsentrasi minimum yang dapat diserap akar, hal ini terjadi sebagai akibat terjadinya proses penyerapan ion fosfat yang ada di permukaan akar. Di bawah konsentrasi minimum tersebut akar tidak mampu lagi menyerap P dan unsur hara lainnya, sedangkan pada akar bermikoriza, penyerapan tetap terjadi sekalipun konsentrasi ion fosfat berada di bawah konsentrasi minimum yang dapat diserap oleh akar. Proses ini ini terjadi karena afinitas hifa eksternal yang lebih tinggi atau peningkatan daya tarikmenarik ion-ion fosfat yang menyebabkan pergerakan P lebih cepat ke dalam hifa MVA.
JAKARTA, KOMPASINDO.CO.ID, 20 Desember 2023 – Dalam sebuah wawancara eksklusif di Sawit Indonesia Award 2023 yang diselenggarakan di Hotel Borobudur Jakarta, John, General Manager PT. Myco Agro Lestari, dengan bangga mewakili merek mereka, MycoVir. Acara penghargaan bergengsi ini mengakui MycoVir sebagai Top 3 Protection Product From Ganoderma, diserahkan oleh Qayuum Amri, Pimpinan Redaksi Majalah Sawit Indonesia, sebagai pengakuan atas kontribusi signifikan mereka untuk kemajuan industri kelapa sawit di Indonesia.
Dalam ungkapannya atas penghargaan kedua kalinya, John menyoroti dedikasi perusahaan terhadap bidang tersebut dan komitmennya terhadap keunggulan. Dia menjelaskan bahwa MycoVir, produk dari PT. Myco Agro Lestari, mengkhususkan diri dalam produksi fungi mikoriza. Jamur ini memainkan peran penting dalam meningkatkan kesehatan tanaman, meningkatkan penyerapan nutrisi, dan berkontribusi pada kesuburan tanah secara keseluruhan.
John menyampaikan, “Komitmen kami tidak hanya melibatkan perlindungan terhadap tanah utama; ini meluas untuk membina sistem akar yang lebih sehat, memastikan tanaman tidak hanya terlindungi tetapi berkembang. Penghargaan terkini kami untuk Top 3 Proteksi Ganoderma adalah bukti dedikasi kami terhadap inovasi dan keunggulan.”
MycoVir Merevolusi Pertanian dengan Pupuk Berkualitas Tinggi yang Memenangkan Penghargaan
Dalam upaya merevolusi pertanian, PT. Myco Agro Lestari memperkenalkan MycoVir, pupuk inovatif dengan formula fungi mikoriza yang kaya. Diakui karena kemampuan infeksinya yang luar biasa hingga 90%, MycoVir bertujuan mengubah perkebunan menjadi tanah subur. Dampak pupuk ini tidak hanya pada kesehatan tanaman, tetapi juga menyerap karbon sebanyak 13,12 gigaton, setara dengan mengurangi 36% emisi bahan bakar fosil global tahunan.
Keunggulan Utama Pupuk MycoVir:
Peningkatan produktivitas tanaman Peningkatan kualitas tanah Peningkatan biomassa akar untuk penyerapan hara maksimal Petani diundang untuk mengunjungi www.mycovir.com untuk informasi lebih lanjut dan memesan pupuk inovatif ini secara daring.
Optimalkan Hasil Pertanian Hortikultura dengan Pupuk MycoVir GOLD
MycoVir GOLD membawa pertanian hortikultura ke tingkat baru dengan pertumbuhan dan kesehatan maksimal. Mengaktifkan fosfat tanah dan melindungi dari serangan jamur patogen tular tanah, MycoVir GOLD menjamin keseimbangan ekosistem mikroorganisme tanah.
Keunggulan Utama Pupuk MycoVir GOLD:
Stimulasi pertumbuhan yang cepat Peningkatan kualitas panen Keseimbangan mikroorganisme tanah Tingkatkan potensi penuh setiap tanaman dalam usaha hortikultura Anda. Temukan lebih lanjut tentang MycoVir GOLD di www.mycovirgold.com dan pesan pupuk ini sekarang juga.
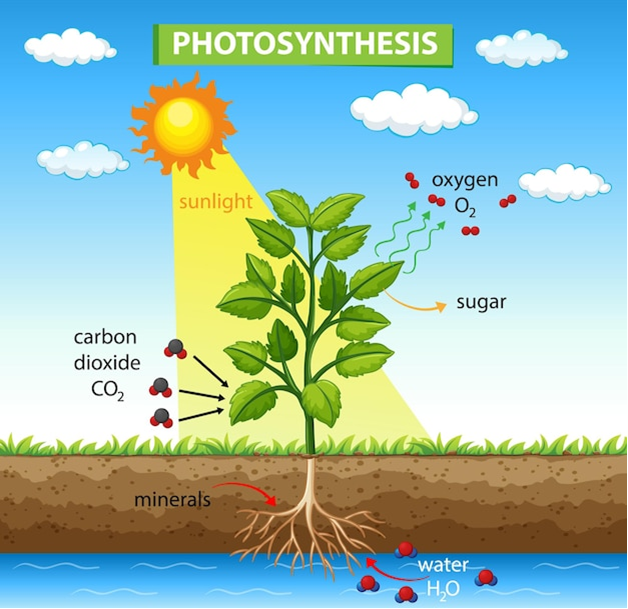
Fotosintesis adalah proses biokimia penting & mendasar yang memungkinkan tanaman mengubah energi cahaya menjadi energi kimia, mendorong pertumbuhannya dan berkontribusi terhadap perkembangan ekosistem yang kita hargai. Proses Fotosintesis: Fotosintesis terjadi terutama di kloroplas sel tumbuhan, di mana pigmen khusus, terutama klorofil, memanfaatkan energi pancaran sinar matahari. Energi ini kemudian dimanfaatkan untuk mengubah karbon dioksida (CO2) dan air (H2O) menjadi glukosa (C6H1206) dan oksigen (02).
Penyerapan Cahaya: Pigmen klorofil menyerap cahaya terutama pada panjang gelombang biru dan merah, sehingga memicu serangkaian reaksi di dalam kloroplas.
Pemisahan Air: Energi cahaya memulai pembelahan molekul air (fotolisis) menjadi oksigen, proton, dan elektron. Oksigen dilepaskan sebagai produk sampingan, berkontribusi terhadap oksigen yang kita hirup.
Fiksasi Karbon Dioksida: Energi yang ditangkap menggerakkan siklus Calvin, serangkaian reaksi enzimatik yang menggunakan elektron dan proton yang dilepaskan untuk mengubah CO2 dari atmosfer menjadi karbohidrat, khususnya glukosa, yang berfungsi sebagai sumber energi utama bagi tanaman.
Signifikansi dalam Pertumbuhan Tanaman: Fotosintesis adalah landasan pertumbuhan dan perkembangan tanaman. Ini adalah proses utama yang bertanggung jawab untuk mensintesis senyawa organik, menyediakan energi yang diperlukan untuk aktivitas seluler, dan memfasilitasi pembentukan biomassa. Proses ini sangat penting dalam:
Produksi Energi: Menghasilkan glukosa dan gula lainnya sebagai sumber energi bagi tanaman.
Pembentukan Biomassa: Membantu pembentukan komponen struktural dan membantu pertumbuhan.
Pelepasan Oksigen: Penting untuk respirasi tumbuhan dan organisme hidup lainnya.
Faktor Penting untuk Fotosintesis:
Cahaya: Jumlah sinar matahari yang optimal sangatlah penting. Cahaya yang terlalu sedikit menghambat proses, sedangkan intensitas cahaya yang berlebihan dapat merusak tanaman.
Karbon Dioksida: Tingkat CO2 yang cukup sangat penting agar siklus Calvin dapat berjalan secara efisien.
Air: Ketersediaan air yang cukup mendukung fotolisis, menjamin pasokan elektron dan proton yang berkelanjutan.
Suhu: Fotosintesis beroperasi secara optimal dalam kisaran suhu tertentu. Panas atau dingin yang ekstrim dapat menghambat proses tersebut.
Klorofil: Kehadiran klorofil dan pigmen lain memungkinkan penyerapan cahaya, yang memulai proses tersebut.
Fotosintesis merupakan bukti keselarasan rumit antara tumbuhan dan lingkungannya. Pentingnya hal ini dalam mendorong pertumbuhan tanaman dan mempertahankan kehidupan di Bumi tidak dapat dilebih-lebihkan. Memahami proses dan faktor-faktor yang mempengaruhinya akan memperjelas keseimbangan yang diperlukan untuk kemakmuran dunia hijau kita.
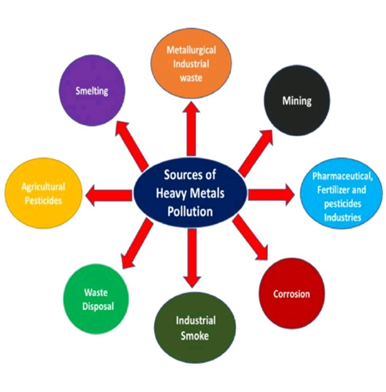
Tanah merupakan komponen penting dari lingkungan yang kesuburannya dapat dipengaruhi oleh keberadaan logam berat. Akumulasi logam berat di dalam tanah akan menurunkan kualitas kesuburan tanah dan membahayakan ketahanan pangan serta kesehatan manusia dan organisme lainnya. Hal ini juga mempengaruhi jumlah keanekaragaman dan aktivitas mikroba tanah sehingga produksi enzim ekstraseluler berkurang. Enzim tanah lebih sensitif terhadap cekaman logam berat dibandingkan tumbuhan dan hewan. Dengan demikian, aktivitas enzim tanah dapat menjadi indikator yang mencerminkan kondisi dan kualitas tanah.
Perkembangan masyarakat dan aktivitas industri telah meningkatkan jumlah logam berat yang masuk ke dalam tanah. Berbeda dengan polutan organik, logam berat tidak dapat terurai di lingkungan alamiah Distribusi, mobilitas, bio-availabilitas, dan toksisitas logam-logam ini di dalam tanah ditentukan oleh unsur kimianya. Logam berat dapat terperangkap dalam struktur bangun mineral silikat atau terikat pada berbagai senyawa tanah, termasuk bahan organik, mineral lempung, oksida besi dan mangan serta mineral karbonat. Daya ikat logam berat dengan berbagai senyawa beragam ini mempengaruhi penyimpanan atau pelepasannya dari dalam tanah.
Bentuk kimia logam berat mempengaruhi kelarutan, ketersediaan dan toksisitas logam tersebut di dalam tanah. Oleh karena itu, untuk memperkirakan pengurangan efek-efek tersebut akibat penerapan adsorben yang berbeda maka bentuk kimia logam dalam tanah harus ditentukan lebih dulu. Metode ekstraksi sekuensial digunakan untuk menentukan bentuk kimia logam berat dan bio-availabilitasnya. Sebaran bentuk kimia logam dalam tanah bervariasi dan bergantung pada pH, kapasitas tukar kation, jumlah bahan organik, tekstur tanah, potensial oksidasi dan reduksi, jumlah kapur dan jumlah oksida Fe-Mn.
Memperbaiki kondisi tanah yang terkontaminasi logam berat dengan metode konvensional seperti penggalian, pencucian asam, ekstraksi dan stabilisasi spesies ionik menggunakan elektroda sangat mahal dan memakan waktu. Sebaliknya metode imobilisasi kimia logam berat dalam tanah merupakan teknologi yang hemat biaya dan mudah digunakan saat ini untuk memulihkan tanah yang terkontaminasi. Metode imobilisasi kimia didasarkan pada pengurangan bentuk logam yang tersedia di dalam tanah dengan menggunakan berbagai bahan tambahan seperti bahan organik, oksida besi dan mangan, mineral tanah liat dan polimer. Oleh karena itu, ketersediaan logam dalam tanah berkurang karena adanya ikatan dengan adsorben. Struktur morfologi dan kimia dari adsorben dan luas permukaannya mempunyai pengaruh terhadap sifat adsorpsinya.
Beberapa adsorben yang mempunyai pengaruh tinggi terhadap stabilisasi logam berat dalam tanah: Mineral lempung seperti: Bentonit, Diatomit, Zeolit. Karbon Aktif, Kitosan, Fe dan Mn oksida, Biochar, Senyawa organik, Komposit nano.
Pertumbuhan dan perkembangan tanaman sangat bergantung pada kecukupan dan konsentrasi unsur hara mineral yang tersedia di dalam tanah. Tanaman sering kali menghadapi tantangan yang signifikan untuk memperoleh pasokan nutrisi yang cukup untuk memenuhi kebutuhan proses seluler dasar karena imobilitas relatifnya. Kekurangan dari salah satu unsur hara tersebut dapat mengakibatkan penurunan produktifitas dan/atau kesuburan tanaman. Gejala kekurangan unsur hara seperti pertumbuhan terhambat, kematian jaringan tanaman, atau daun menguning karena berkurangnya produksi klorofil, pigmen yang diperlukan untuk fotosintesis.
Defisiensi unsur hara berdampak signifikan terhadap tanaman, dapat mengakibatkan berkurangnya hasil panen atau menurunnya kualitas tanaman. Defisiensi ini juga dapat menyebabkan berkurangnya keanekaragaman hayati secara keseluruhan karena tanaman berperan sebagai produsen yang mendukung sebagian besar jaringan makanan. Unsur hara mineral biasanya diperoleh dari tanah melalui akar tanaman, namun banyak faktor yang dapat mempengaruhi efisiensi perolehan unsur hara. Faktor-faktor tersebut adalah:
Kerusakan akibat hama seperti: cacing akar putih, agrotis, penyakit akar dan hama menghambat penyerapan unsur hara dengan mengubah bentuk akar dan mengganggu aktifitas fisiologi akar
Kerusakan akibat hama seperti: cacing akar putih, agrotis, penyakit akar dan hama menghambat penyerapan unsur hara dengan mengubah bentuk akar dan mengganggu aktifitas fisiologi akar
Suhu tanah rendah. Suhu tanah juga mempengaruhi penyerapan unsur hara dengan mengubah viskositas air tanah dan transportasi unsur hara oleh akar. Pada suhu yang rendah, penyerapan unsur hara oleh tanaman berkurang akibat tingginya viskositas air tanah dan rendahnya aktifitas pengangkutan unsur hara oleh akar.
Tekstur tanah. Tekstur tanah mempengaruhi kararkterisitik tanah berikut:
kapasitas menahan air tanah
kapasitas menahan unsur hara
erodibilitas
kemampuan kerja penetrasi akar
porositas
Tekstur tanah mempengaruhi kesuburan tanah dan ketersediaan unsur hara.
Mayoritas kekurangan sulfur terjadi di tanah berpasir.
Nitrogen mudah tercuci di tanah berpasir. Hilangnya nitrogen tanah (denitrifikasi) lebih sering terjadi pada tanah liat berat.
Kalium dapat larut di tanah berpasir tetapi tidak dapat bergerak di tanah bertekstur sedang hingga halus.
Kekeringan. Penurunan penyerapan unsur hara selama kekeringan dapat terjadi karena beberapa alasan, termasuk berkurangnya pasokan unsur hara akibat berkurangnya mineralisasi dan berkurangnya difusi dan aliran massa unsur hara di dalam tanah
Pemadatan tanah. Pemadatan tanah berarti meningkatnya kepadatan tanah. Akibatnya akar kurang mampu menembus tanah dan umumnya dangkal serta keras. Dengan pertumbuhannya yang terbatas, akar kurang mampu memanfaatkan tanah untuk mendapatkan nutrisi dan kelembabaan. Dalam situasi ini defisiensi nitrogen dan kalium adalah hal yang paling umum terjadi
Kurangnya bahan organik tanah. Hilangnya kandungan karbon organik tanah akan membatasi kemampuan tanah dalam menyediakan nutrisi untuk produksi hasil tanaman yang berkelanjutan. Kurangnya karbon organik juga berarti lebih sedikit makanan bagi organisme hidup yang ada di dalam tanah, sehingga mengurangi keanekaragaman hayati di dalam tanah.
Abstract Basal stem rot of oil palm caused by Ganoderma boninense is the most serious disease of oil palm in Malaysia, Indonesia, and other oil-palm-producing countries. Economic losses caused by the disease can be up to USD500 million a year. For many years, basal stem rot was found to infect older palm trees of more than 25 to 30 years in age. Only in the 1950s, the disease began to appear in much younger palm trees, 10 to 15 years old, and, in the last decade or so, palm trees as young as 1 year were infected by the disease. The highest incidence occurs in coastal areas of Southeast Asia but the disease has now infected oil palm in inland areas, mainly oil palm planted in peat soils. Disease incidence is also high in areas previously growing coconut or forest. Basal stem rot infection and spread occur through root-to-root contact, and basidiospores that colonize the roots also play a role. In the early stages of infection by G. boninense, the pathogen behaves as a biotroph and later as a necrotroph, secreting cell-wall-degrading enzymes and triggering host defense responses. Genes, gene products, and metabolic pathways involved in oil palm defense mechanisms against G. boninense have been identified and these metabolites have the potential to be used as markers for early detection of the disease. Integrated disease management used to control basal stem rot includes cultural practices, chemical control, and application of biocontrol agents or fertilizers. Early detection tools have also been developed that could assist in management of basal stem rot infections. Development of resistant or tolerant oil palm is still at an early stage; therefore, the existing integrated disease management practices remain the most appropriate methods for managing basal stem rot of oil palm.
Oil palm (Elaeis guineensis Jacq.) is grown in tropical regions of Asia, Africa, and South and Central America that receive high rainfall. The major producing areas are located between the latitudes 7°N and 7°S. Oil palm is the second major vegetable oil crop after soybean and constitutes approximately 40% of all commercially produced vegetable oils (Murphy et al. 2021). Oil palm is regarded as the most space-efficient vegetable oil crop, because 1 ha of trees can produce up to eight times more oil than other oil crops (Zulkifli et al. 2010). Globally, annual palm oil production is approximately 81 million tons from approximately 19 million ha of planting area (Murphy et al. 2021). Palm oil is used to manufacture various edible and nonedible products, including food, cosmetics, biofuels, detergents, plastics, and industrial chemicals (Murphy et al. 2021).
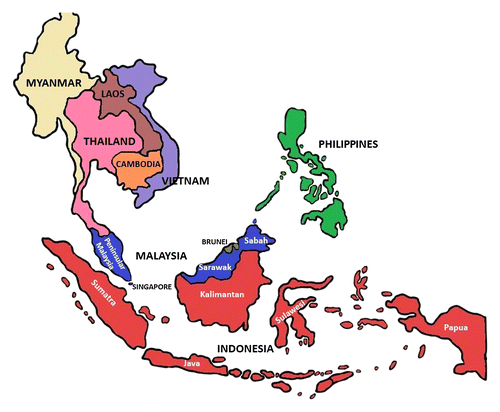
Oil palm trees thrive in hot and humid conditions, which are common in Southeast Asian regions. The first oil palm plantation was established in Peninsular Malaysia (also known as West Malaysia) in 1917; large-scale cultivation began in 1961 owing to the Malaysian government’s encouragement for agricultural diversification to reduce dependence on natural rubber (Hartley 1977). Over the years, oil palm plantations have expanded to East Malaysia in the states of Sabah and Sarawak. In Indonesia, oil palm plantations are concentrated on the islands of Sumatra and Kalimantan (Fig. 1) (Paterson 2020). Malaysia and Indonesia have dominated the palm oil trade since 1966 and currently account for over 84% of the global palm oil production, with Indonesia being the largest producer (Statista 2020). Approximately 60% of palm oil is exported to India, Pakistan, Bangladesh, China, and European Union countries (Statista 2020).
Fig. 1. Map of Southeast Asia region and countries.
Apart from Malaysia and Indonesia, oil palm is grown in at least 43 other countries (Fig. 2), including Thailand, the Philippines, and Myanmar; in the latter two countries, oil palm plantations are at an early stage of development (Corley and Tinker 2003; Murphy et al. 2021). In Africa, major oil-palm-cultivating countries are Nigeria, Ghana, Côte d’Ivoire, Cameroon, Sierra Leone, Benin, Angola, and the Democratic Republic of Congo. Only Cameroon and Côte d’Ivoire produce palm oil for export, whereas the other African countries use it for local consumption (Paterson 2021). Currently, Colombia and Ecuador are the primary oil palm producers in the Americas, although the oil palm industry in Brazil is also expanding (Murphy 2014; Nahum et al. 2020).
One of the major disease concerns associated with large-scale oil palm plantations in Malaysia and Indonesia is basal stem rot (BSR), caused by the white rot fungus Ganoderma boninense. The problem is especially acute in replanted plantations; disease incidence has increased over the decades due to repeated replanting, and can reduce yield by 50 to 80% (Paterson 2019).
Early studies on BSR and Ganoderma spp. were conducted in the 1960s, particularly in Malaysia, when information related to their incidence, infection, spread, treatment, and control methods was published (Navaratnam 1964; Tan 1968; Turner 1965a, b, c, d; Turner and Bull 1967). Research published in the 1980s through early 2000s focused on pathogen species identification, pathogenicity, and pathogen mating systems, as well as quantifying incidence and spread of BSR. Characterization studies of Ganoderma spp. using molecular methods and host–pathogen interactions were also performed. More data were obtained after the establishment of oil palm plantations in Indonesia and Papua New Guinea.
Although information on the cause of infection and spread of BSR has been well documented, development of early detection methods for this disease is still ongoing. This review presents an overview of recent progress in improving detection methods and disease management, and offers possible future directions for BSR research.
Oil Palm
Oil palm (E. guineensis Jacq.) originated in West Africa, particularly in the tropical forests of Côte d’Ivoire, Nigeria, Ghana, Cameroon, Democratic Republic of Congo, and Angola (Hartley 1988). In African countries, palm oil has long been used as a primary ingredient in several traditional dishes (Poku 2002).
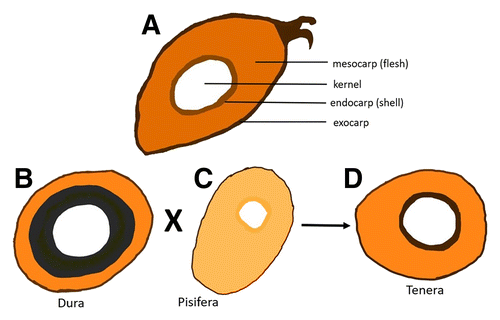
Oil palm is a monocot in the family Arecaceae (Palmae), which bears both male and female inflorescences separately on the same tree (Hartley 1988). Two major species of oil palm are E. guineensis, known as African oil palm (Fig. 3A and B), and E. oleifera, the American oil palm, found in South and Central America. E. guineensis is widely planted commercially because of its high oil content. E. oleifera is not planted for commercial purposes but it contains sources of genetic resistance against BSR, which is useful in breeding programs (Durand-Gasselin et al. 2005).
Fig. 3. African oil palm (Elaeis guineensis). A, Mature tree. B, Young tree.
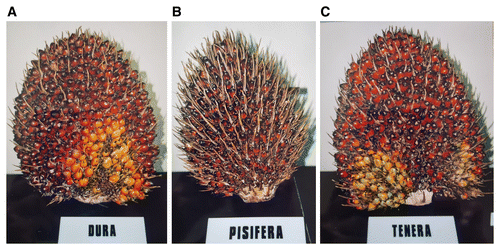
The three recognized cultivars of E. guineensis are Dura, Tenera, and Pisifera, categorized according to the shell thickness or endocarp and mesocarp contents (Fig. 4) (Corley and Tinker 2003; Latiff 2000). Dura has a large kernel, a thick shell (Fig. 4A), large bunches of fruit, and a high oil yield, whereas Pisifera is shell-less and has a small kernel, large mesocarp, and poor oil yield owing to high female infertility; it is mainly used in breeding (Fig. 4B). Tenera is a hybrid of Dura and Pisifera (Fig. 4C), with a good kernel size, a thin shell that makes processing easier, a large mesocarp, and an oil yield 25 to 30% higher than that of Dura fruit (Corley and Tinker 2003). Two types of palm oil are produced by E. guineensis: from the fibrous mesocarp and from the palm kernel (Corley and Tinker 2003). Fruit bunches of Dura, Pisifera, and Tenera are shown in Figure 5.
Fig. 4. Cross section of three oil palm cultivars. A, General structure of oil palm fruit; B, Dura (thick shell); C, Pisifera (shell-less); and D, Tenera (less thick shell).
Fig. 5. Oil palm fruit bunches of cultivars A, Dura; B, Pisifera; and C, Tenera. Photos taken at United Plantation, Teluk Intan, Perak, Malaysia.
Oil palm fruit can be harvested after 2 to 3 years of planting, although the productive age is 9 to 15 years. Oil palm has a life span of 25 to 30 years, after which the tree becomes too tall for harvesting the fruit bunches. In Malaysia, some long-established oil palm plantations are already in their third generation (Basiron 2007).
Pathogen
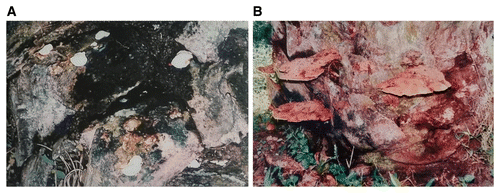
Several species of Ganoderma have been reported to be associated with BSR. Initially, G. lucidum was reported as the causal pathogen but later it was found that G. lucidum is limited to temperate regions (Steyaert 1972). Six species of Ganoderma, G. boninense, G. miniatocinctum, G. chalceum, G. tornatum, G. zonatum, and G. xylonoides, either saprophytic or parasitic on oil palm, were identified based on their fruiting bodies and basidiospore characteristics (Steyaert 1972). The basidiocarp of G. boninense at the base of an oil palm tree and a bracket-shaped basidiocarp of the fungus are shown in Figure 6.
Fig. 6. Basidiocarp of Ganoderma boninense. A, Basidiocarp at the base of an oil palm tree. B, Spore deposits on the upper part of basidiocarp. C and D, Reddish-brown basidiocarp.
In Peninsular Malaysia, Ho and Nawawi (1985) found that G. boninense was the most common species associated with BSR. G. boninense was later confirmed, using pathogenicity tests, to be the causal agent of BSR (Khairudin 1990). Furthermore, G. boninense has been reported as a causal pathogen of BSR in Sabah and Sarawak, Malaysia (Chong et al. 2011; Midot et al. 2019), Indonesia, and Papua New Guinea (Haryadi et al. 2019; Pilotti et al. 2004; Purba et al. 2019). In addition to oil palm, G. boninense also causes BSR in several other palm trees, including foxtail palm (Wodyetia bifurcata), which is a landscape palm (Mohd Farid et al. 2018); coconut (Snehalatharani et al. 2016); and the ornamental palm Chrysalidocarpus lutescens (Lim and Fong 2005).
G. boninense isolates from Malaysia, Sumatra, Indonesia, and Papua New Guinea originating from the same oil palm plantation, as well as from the same individual tree, were genetically variable (Latiffah et al. 2005; Mercière et al. 2017; Miller et al. 1999; Pilotti et al. 2003; Wong et al. 2021). High levels of genetic variation in G. boninense isolates may be associated with the ability of G. boninense isolates to adapt to various substrates such as soil, plant debris, and new oil palm stands (Latiffah et al. 2005). Genetic variation of G. boninense also may be associated with heterothallism and the tetrapolar mating system of the pathogen, with multiple alleles occurring at both mating type loci, which favors outcrossing (Sanderson et al. 2000).
More information on the population structure and genetic variation of G. boninense in Malaysia and Indonesia was obtained using microsatellite markers. Based on the data from 11 microsatellite markers, G. boninense isolates from Peninsular Malaysia and Sumatra, Indonesia exhibited a high degree of genetic variation. Genetic differentiation of G. boninense isolates from both areas was not observed, which demonstrated that spores may play a role in spreading the pathogen in these two areas (Mercière et al. 2017).
Similar results were also reported by Wong et al. (2021) using 16 polymorphic microsatellite markers, among which G. boninense in the Peninsular Malaysia, in Sarawak, Malaysia, and in Sumatra, Indonesia, demonstrated a high level of genetic variation. High genetic variation was observed among isolates within a population as well as among different populations due to the outcrossing of the pathogen’s basidiospores. High gene flow was detected among G. boninense populations but genetic differentiation was not observed, which indicated that the BSR pathogen populations in the three oil palm planting areas were similar. Moreover, three admixed genetic clusters of the pathogen population were detected in Peninsular Malaysia, in Sarawak, Malaysia, and in Sumatra, Indonesia (Wong et al. 2021). Based on mating studies, internal transcribed spacer (ITS) sequences, and microsatellite genotyping, conspecificity of G. boninense isolates from four oil-palm-producing countries (Indonesia, Malaysia, Papua New Guinea, and Solomon Islands) was demonstrated (Pilotti et al. 2021).
In contrast to the high genetic variation reported in a few studies, low genetic variation was observed in G. boninense isolates collected from eight locations of coastal areas in the state of Sarawak, Malaysia. Low genetic variation was detected in 117 isolates using sequences from the ITS region (Midot et al. 2019). The low genetic variation might be because the plantation was a first-generation planting and was previously a secondary forest, which may be associated with low disease pressure. G. boninense isolates in first-generation plantations are regarded as the founder population and are later adapted to oil palm as the host (Midot et al. 2019).
Other species of Ganoderma reported from BSR-infected oil palm trees are G. miniatocinctum, G. chalceum, and G. zonatum. Pathogenicity tests revealed that these species are pathogenic and cause BSR in oil palm (Idris et al. 2000). In Sarawak, Malaysia, G. zonatum and G. miniatocinctum are also associated with BSR (Wong et al. 2012). In Papua New Guinea, G. tornatum was found on infected oil palm trees and considered a minor pathogen (Pilotti et al. 2004). G. zonatum was identified as the causal pathogen of BSR in Colombia (Castillo et al. 2022).
BSR of oil palm has also been reported in southwestern Cameroon, where G. ryvardense was found to be associated with the disease (Kinge and Mih 2011). According to Wang et al. (2014), G. ryvardense is closely related to G. boninense, and they may be synonymous. G. ryvardense may also be conspecific to the BSR pathogen in Malaysia and Papua New Guinea (Pilotti et al. 2021). Another causal pathogen of BSR reported in Cameroon is G. lobenense (Kinge and Mih 2014). Further studies such as mating compatibility tests and molecular genetic analysis may help to clarify the identity and confirm the conspecificity of G. ryvardense with G. boninense as well as its relatedness with G. lobenense.
G. boninense is heterothallic with bifactorial incompatibility due to two nonlinked loci, mating-type A and mating-type B (Pilotti et al. 2002). Mating tests indicated that G. boninense is compatible with G. miniatocinctum and G. zonatum as well as Ganoderma isolates from two ornamental palm trees: sealing wax palm and McArthur palm. Mating test results indicated that the alleles involved in mating occurred between different mating types, resulting in formation of 100% fertile dikaryons. The findings suggested that G. boninense, G. zonatum, and the other isolates could belong to the same biological species, and cross infection could occur among Ganoderma spp. and between the ornamental palm and oil palm (Chan et al. 2015).
Occurrence of BSR in the Oil Palm
BSR has been reported in almost all palm-oil-producing countries. The incidence of serious diseases has increased over the years as replanting of oil palm occurs, particularly in Malaysia and Indonesia. In some plantations, replanting of the crops is in the third generation, which makes the palm trees more susceptible to BSR infection due to inoculum buildup.
Incidence of BSR is much more serious in Peninsular Malaysia than in the states of Sabah and Sarawak in East Malaysia. The higher BSR incidence in Peninsular Malaysia might be due to earlier large-scale cultivation of oil palm and high occurrence of replanting. BSR can be found in oil palm trees as young as 1 year old (Ariffin et al. 1989a; Turner 1981). In Indonesia, a higher BSR incidence was reported in Sumatra and Java than in Kalimantan, Sulawesi, and Papua, probably due to younger plantings in the former regions (Suryantini and Wilandari 2018). In North Sumatra, after 25 years, 40 to 50% of the oil palm trees were infected (Subagio and Foster 2003).
Although incidence of BSR in Papua New Guinea is not as high as in Malaysia and Indonesia, 50% disease incidence has been reported in some oil palm plantations there. According to Murphy et al. (2021), the incidence rate of BSR infection in Papua New Guinea is likely to be 25% on average, considerably lower than in Malaysia and Indonesia. BSR incidence is relatively low in oil palm plantations in Thailand, where the disease mainly affects older palm trees and is more widespread and serious in the southern part of the country (Pornsuriya et al. 2013).
Colombia is the largest oil-palm-producing country in the Americas, with a low BSR incidence rate. However, in the last decade, BSR has emerged as a growing concern, posing a serious threat to the oil palm industry there (Arango et al. 2016; Martínez 2011).
In Africa, BSR has been reported in Nigeria, Ghana, Democratic Republic of the Congo, Angola, Tanzania, North Mozambique, and Cameroon (Turner 1981). However, recent information on the occurrence of BSR in sub-Saharan Africa is lacking, probably due to the prevalence of Fusarium wilt in the region. The only available information on the occurrence of the disease is in Cameroon. In southwestern Cameroon, BSR incidence was higher in younger oil palm trees (16 years old or younger) than in older ones (more than 30 years old). In general, BSR incidence in oil palm trees of the same age in Cameroon was lower than that in Malaysia and Indonesia (Mih and Kinge 2015).
In Malaysia, BSR was first reported by Thompson (1931) in oil palm that was over 25 years old, and which was due for replanting. For several years, BSR was considered to be a disease of older palm trees. It was only in the late 1950s that it began to appear in much younger palm trees (10 to 15 years old) and, in the last three decades, palm trees as young as 1 year old were found to be infected (Ariffin et al. 1989b; Turner 1981). Infected young palm trees commonly die within 6 to 24 months after initial symptoms appear, whereas mature palm trees can survive for 2 to 3 years or more after infection (Corley and Tinker 2003). Similar BSR occurrence was also observed in North Sumatra, Indonesia. The disease was found to infect younger palm trees in a first-generation planting. The survey also indicated that BSR incidence was higher in younger palm trees, which was attributed to existing inoculum in the field (Lisnawita et al. 2016; Susanto et al. 2008; Treu 1998).
The highest BSR incidence is observed in coastal areas (Khairudin 1990; Turner 1981). This is attributed to the prevalence of plantings in previous coconut stands and the nature of the soil and its water table. Coastal soils are mainly clay, silty clay, or clay loam. These types of soils with poor internal drainage and a high water-retention capacity are likely favorable for the growth of Ganoderma spp., since the fungus prefers high moisture conditions (Singh 1990; Turner 1981).
The disease is severe in areas where old coconut trees are felled and their stumps are buried or left on the ground to rot. Disease incidence is also high when old coconut trees are poisoned and left standing to rot in situ. The coconut stumps that are left on the ground to rot become a substrate for formation of G. boninense fruiting bodies (Turner 1981).
A recent study by Mohd Shukri et al. (2020) on the incidence of BSR in oil palm plantations owned by smallholders indicated that, regardless of the soil type, oil palm is susceptible to BSR infection. Highest disease incidence was detected in lateritic soil (27.7%), followed by coastal (14%), inland (8.72%), and peat (6%) soils. Earlier, it was predicted that peat soil is not favorable for BSR incidence but that claim has now been disproved (Ariffin et al. 1989b; Cooper et al. 2011). BSR has also infected oil palm plantations grown on peat soil in Indonesia, and high incidence of the disease has been observed (Susanto and Huan 2010). A study on BSR incidence in peat soil plantations that was conducted in North Sumatra and West Kalimantan, Indonesia suggested that high incidence of BSR in peat soil may be due to several factors (Supriyanto et al. 2020; Susanto et al. 2008). G. boninense survives as a saprophyte and has adapted to the conditions of the peat soil environment. Monocultures that eliminate other woody plants may increase likelihood of oil palm infection (Supriyanto et al. 2020). In addition to tree susceptibility, selective pressure on the pathogen may also play a role due to changes of the ecosystem (Lucas et al. 2015). G. boninense infection may also be associated with deficiencies of copper and zinc, which are common in the very-low-pH conditions of peat soil (Supriyanto et al. 2020).
Infection Process
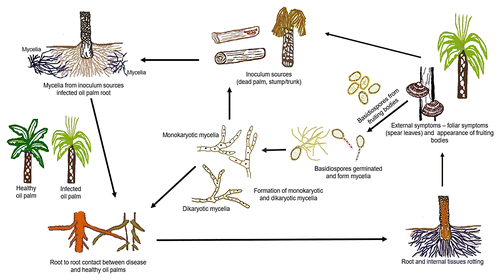
Spread of BSR to healthy palm trees is associated with root contact with a source of inoculum, including stumps, trunks, and infected oil palm trees (Fig. 7). A high disease incidence has been observed in areas where the trunks and stumps of coconut colonized by G. boninense act as inoculum. The pathogen enters the roots and then spreads to the stem base (Fig. 7) (Ho and Khairudin 1995; Turner 1981). In a field trial by Rees et al. (2007), inoculum (Ganoderma-infested palm or rubber wood blocks) was required to infect over the course of 6 to 8 months.
Fig. 7. Disease cycle of basal stem rot (BSR) of oil palm.
Infection occurs when mycelia penetrate the root and spread to the cortex and stem base. In the pathogen’s biotrophic phase, numerous intracellular hyphae invade and occupy the host roots and stem base tissues. At this stage, the oil palm tissues remain intact. Later, the pathogen switches to necrotrophy, with extensive cell wall degradation. The third phase is the formation of pseudosclerotia embedded in extensive and tough mycelia surrounding the roots (Rees et al. 2009).
During root colonization, the pathogen secretes cell-wall-degrading enzymes to degrade root components, including lignin and cellulose (Cooper 1984). The breakdown of lignin results in bleaching, which is indicative of the white rot phase of the disease (Paterson 2007). Colonization of the root cortex and stem parenchyma produces holes in cell wall layers, leading to cell wall degradation. During infection, starch can be used by the pathogen as a carbon source (Rees 2006). Infected oil palm trees are not easy to diagnose at this stage, except for external symptoms such as unopened spear leaves (the youngest emerging leaves) and the appearance of fruiting bodies at the base of the trunk (Figs. 8 and 9).
Fig. 8. External symptoms of basal stem rot. A and B, Infected oil palm showing formation of spear leaves and skirting (frond hanging downward).
Fig. 9. External symptoms of basal stem rot. Formation of A, white buttons and B, basidiocarp at the base of oil palm tree.
Role of basidiospores
Basidiospores can colonize felled oil palm trees as well as stumps and trunks in the oil palm field, and these substrates become infection foci (Turner 1981). Genetic diversity studies of G. boninense isolates associated with BSR showed high levels of variability, implying that infection could be caused by basidiospores (Latiffah et al. 2005; Mercière et al. 2017; Miller et al. 1999; Pilotti et al. 2021; Wong et al. 2021). Pilotti (2005) detected multiple isolates in a single oil palm, which clearly demonstrated multiple infections and suggested basidiospores as the source of the infection. Infection by means of root-to-root contact may occur relatively infrequently (Pilotti et al. 2018).
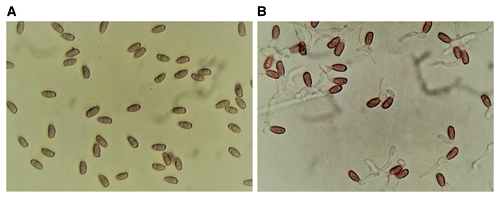
In oil palm plantations, basidiospore germination often occurs at 25 to 31°C (Ho and Nawawi 1986). Nongerminated and germinated basidiospores of G. boninense are shown in Figure 10. At this early stage, monokaryotic mycelia are saprophytic. Germination leads to the formation of monokaryotic mycelia and colonization of the available substrates in the field but this stage is not infectious (Rees et al. 2007). When monokaryotic mycelia encounter compatible monokaryotic mycelia, dikaryotic mycelia are formed, which can cause BSR infection (Chan et al. 2011).
Fig. 10. Basidiospores of Ganoderma boninense. A, Nongerminated spores and B, germinated spores.
Basidiospores can germinate on the cut surfaces of fronds, peduncles, and stems, which indicates their ability to germinate and grow on different parts of oil palm (Rees et al. 2012). G. boninense has also been detected in the rachis, bole, and roots of asymptomatic seedlings, and root colonization by basidiospores and dikaryons is common (Pilotti et al. 2018). Molecular studies revealed highly diverse G. boninense isolates from oil-palm-producing regions, supporting the hypothesis that basidiospores play major roles in BSR infection and spread (Pilotti et al. 2018, 2021; Wong et al. 2021).
BSR symptoms
Initially, an excessive number of spear leaves are observed (Fig. 6A and 6). In healthy plants, only two to three spear leaves are produced per month. The foliage color of infected palm trees is generally much paler green than on healthy palm. Eventually, the fronds die; necrosis begins in the oldest frond and spreads to the younger fronds. In most cases, the dead fronds either droop at the base or fracture at some point along the rachis and hang down to encircle the trunk (Fig. 6B). Ultimately, the trunk fractures at the base and the tree falls. However, the bole, which still contains diseased tissue, remains in the ground (Turner 1981).
A reliable disease symptom is the appearance of lesions at the stem base. The lesion appears as a light brown area of rotting tissues marked by irregular dark zones with an outer edge of irregular yellow zones. The dark zone is called the reaction zone (Turner 1981) or black line (Ariffin et al. 1989a) and consists of masses of swollen hyphal cells that form resting structures. Roots of infected palm trees usually become friable and easily disintegrate (Ariffin et al. 1989a; Turner 1981).
G. boninense basidiomata may or may not develop before the appearance of foliar symptoms. The basidiocarp emerges as white buttons or white small fungal swellings that develop into bracket-shaped fruiting bodies (Fig. 9A). The basidiomata develop at the stem base and occasionally on infected roots near the stem base (Fig. 9B). Eventually the palm dies, either remaining upright or falling over (Turner 1981).
Metabolites and defense-related genes involved in BSR infection
The necrotrophic phase of G. boninense infection results in the secretion of cell-wall-degrading enzymes and triggering of a defense response from the host.
Metabolites that may be involved in defense mechanisms against infection by G. boninense include fatty acids, phenols, lipids, phytohormones, and aromatic compounds. Al-Obaidi et al. (2014) reported several host-derived proteins associated with BSR infection in the root of oil palm such as caffeoyl-CoA O-methyltransferase, caffeic acid O-methyltransferase, enolase, fructokinase, cysteine synthase, malate dehydrogenase, and ATP synthase.
Fatty acids and phenols were detected as defensive compounds in the root extract of infected oil palm. Some of the identified fatty acids and phenols were benzoic acid, hexadecanoic acid, 9,12-octadecadienoic acid (Z,Z), 9-octadecenoic acid (Z), and their respective methyl esters (Rozlianah et al. 2015).
Several steroid compounds have been identified as plant defense metabolites against G. boninense infection. Three main steroids, namely sitosterol, γ-sitosterol, and stigmasterol, were found in the crude root extracts of both infected tolerant and susceptible oil palm progenies. Other compounds detected were stigmast-5-en-3-ol, campesterol, cholestan, and an ergosterol (α-tocopherol and β-tocopherol) (Nusaibah et al. 2011). Other metabolites associated with the early defense mechanism of oil palm included heterocyclic aromatic organic compounds such as pyridine, benzo[h]quinoline, and pyrimidine; lipids such as fucosterol, betasitosterol, γ-sitosterol, palmitic acid, and linoleic acid; and antioxidants such as α-tocopherol and β-tocopherol (Nusaibah et al. 2011). These metabolites were detected in the root extract of infected samples, and their concentrations were found to be higher in resistant than susceptible oil palm progeny.
Jee and Chong (2015) reported that growth of G. boninense in infected oil palm trees can be suppressed by treatment with caffeic acid, syringic acid, or 4-hydroxybenzoic acid in concentrations ranging from 0.1 to 0.4 mg/ml. Benzoic acid and salicylic acid not only suppress infection by G. boninense but also promote oil palm growth (Surendran et al. 2018).
Some of the antifungal activities of the metabolites are produced by interactions between the host and the pathogen. The pathogen activates oil palm defense mechanisms via secretion of elicitors and the plant, in turn, secretes various types of metabolites to inhibit pathogen colonization. These interactions occur before BSR is established in the host.
Defense-related genes involved in G. boninense infection in oil palm have been identified in several studies. Tee et al. (2013) identified several genes associated with oil palm defense mechanisms, including genes associated with plant signal transduction and genes involved in phytoalexin production. These genes were either upregulated to enhance oil palm defense mechanisms against infection by G. boninense or downregulated to allow it to overcome the oil palm defense system.
Ho et al. (2016) studied transcriptomes of inoculated and noninoculated oil palm seedlings to infer molecular responses of oil palm roots against G. boninense infection. The transcriptomes recovered from inoculated roots included biochemicals involved in the initial stage of infection, including receptors (receptor-like kinases and receptor-like proteins) and cell-wall-degrading enzymes (chitinases and glucanases).
In another transcriptome study, Bahari et al. (2018) identified upregulated and downregulated gene sequences associated with the transition from biotrophic to necrotrophic phases of G. boninense. Differentially expressed genes included defense-related genes (pathogenesis related-proteins, protease inhibitors, and chitinase), the transcription factors EgERF113 and EgMYC2 (regulators of necrotrophic phase), and reactive oxygen species elicitors (peroxidase and NADPH oxidase).
Early detection of BSR
The main aim of early detection of BSR in oil palm tissues is to develop a more effective method to treat the infected palm before the disease spreads further. The earliest methods for BSR detection include laboratory-based methods such as enzyme-linked immunosorbent assay (Shamala et al. 2006) and designing PCR specific primers to detect Ganoderma spp. pathogenic to oil palm (Idris et al. 2010). In addition to laboratory-based methods, there is a rapid progress in the development of methods that use geospatial technologies. Among the geospatial technologies tested, multispectral and hyperspectral remote sensing, terrestrial laser scanning, spatial maps, tomography images, intelligent e-nose and microfocus X-ray fluorescence were found to be able to differentiate between infected and noninfected oil palm (Khosrokhani et al. 2018).
Methods for the early detection of BSR have also been developed in Colombia and Indonesia. Arango et al. (2016) developed electric impedance tomography based on the color spectrum to distinguish between healthy and infected oil palm trees. In Indonesia, satellite images have been used to identify the spatial patterns of BSR infection in oil palm plantations (Santoso et al. 2011). Multispectral aerial photographs from unmanned aerial vehicles, combined with a vegetation index, have also been used to map BSR incidence and estimate the levels of infection (Wiratmoko et al. 2018). Levels of BSR infection have also been detected using an electric nose device (Kresnawaty et al. 2020).
Reliable and rapid early detection of BSR can be used as a complement to other control methods. However, most of these methods are at the preliminary stage and have been tested in only a few plantations. Moreover, some of the methods require specialized equipment or highly skilled workers to collect and interpret the data, which leads to high costs. Currently, only established, large-scale plantations can afford these technologies. Smallholders still depend on conventional methods such as the observation of typical BSR symptoms to detect infection.
Management of BSR
Integrated disease management by combining several control methods is required to manage BSR because there is no single effective control measure that can stop the spread of this disease. Current control measures include chemical fungicides, cultural practices, and biological methods, as well as fertilizer application. The aim of BSR integrated disease management is to minimize the incidence of BSR after replanting.
Fungicides
Since early studies of BSR, fungicides have long been used to treat the oil palm trees infected with G. boninense. For example, organo-mercury formulations and potassium hydroxyquinoline sulfate have been injected into diseased palm trees. However, because of the large mass of many lesions, it is difficult to accurately estimate the extent of trunk lesions. Therefore, trunk injection using these materials has had only limited success (Turner 1981).
Khairudin (1990) reported slowing the spread of BSR by injecting systemic fungicides such as fusilazol, hexaconazole, cyproconazole, flutriafol, triadimenol, tridemorph, and oxycardoxin. Ariffin and Idris (1991) showed that injection of a fumigant fungicide, dazomet, into the stem lesion could inactivate the production of fruiting bodies, which further slowed the spread of BSR. Idris and Maizatul (2012) later reported that dazomet reduced the spread of BSR by killing Ganoderma inoculum, thereby improving the productivity of the oil palm.
Application of hexaconazole via pressure injection, trunk injection, soil drenching, and soil injection into diseased oil palm trees reduced the spread of BSR in the infected oil palm trees (Idris et al. 2002, 2004c). Pressure injection can be used to accurately apply the fungicide to the infected area. Idris et al. (2004c) reported that, after 5 years, 70% of treated oil palm trees could produce fruit bunches.
Pyraclostrobin, a broad-spectrum fungicide with preventive, curative, and long residual properties, was evaluated for its efficacy in inhibiting the growth of G. boninense. Pyraclostrobin was effective in controlling BSR in oil palm seedlings and showed positive physiological effects on seedlings, including increased height, bole diameter, root mass, photosynthesis rate, PSII quantum efficiency, and relative leaf chlorophyll content (Nurfaezah et al. 2019). However, the effectiveness of this fungicide on oil palm trees beyond the seedling stage has not yet been tested.
Nanotechnology tools are also being applied for delivery of fungicides (Maluin et al. 2019, 2020). A nanodelivery system consisting of hexaconazole encapsulated into chitosan nanoparticles as carrier effectively transported the fungicides to G. boninense cells. The nanodelivery system showed low phytotoxicity and high antifungal activity against the pathogen (Maluin et al. 2019).
Despite the availability of some effective fungicides, control methods using fungicides can lead to environmental pollution as well as development of fungicide resistance. For those reasons, alternative control methods such as cultural practices and biological control are preferred.
Cultural practices
Three replanting techniques (underplanting, windrowing, and clean clearing) have been widely used in Malaysian oil palm plantations. Replanting techniques have been reported to have a significant effect on disease incidence in new oil palm stands (Khairudin 1990; Turner 1981).
Underplanting is associated with a relatively high risk of BSR disease. In this method, old coconut palm trees or oil palm trees are poisoned and left to rot, with younger palm trees planted between the older palm trees. Khairudin (1990) showed that 93.0% of the oil palm seedlings grown near diseased stumps were infected.
When a 15-year-old plantation was renovated, the clean clearing technique resulted in 14.0% BSR incidence compared with 33% for underplanting (Khairudin 1990). Similarly, Singh (1990) reported that clean clearing treatment of previous oil palm plantations led to 3.2% disease incidence after 8 years compared with 13.5% for underplanting. In the clean clearing technique, stumps and trunks from previous crops are excavated and split for drying, followed by burning. Plowing of any pieces of Ganoderma-infected tissues from the soil is carried out to prevent them from becoming infection foci (Turner 1981).
The windrowing replanting technique is similar to clean clearing except that the stumps and trunks of previous crops are stacked in the alleys between row without splitting for drying and burning. Disease incidence in oil palm fields that adopt the windrowing replanting technique differed little from that in fields adopting clean clearing (Khairudin 1990, 1993). Thus, of the three replanting techniques, clean clearing and windrowing are safer than underplanting.
In northern Sumatra, Indonesia, impacts of windrowing, fallowing for 1 year, and poisoning on BSR infection were compared for 7 years. Windrowing without poisoning resulted in 41% infection on replanted palm, whereas windrowing with fallowing but without poisoning resulted in 10% infection after 1 year. Windrowing alone led to 28% incidence of BSR. In contrast, windrowing plus poisoning and fallowing led to only a 3% incidence, and fallowing without poisoning resulted in 6% incidence (Virdiana et al. 2010). Based on the study, fallowing of 1 year before transitioning to a new oil palm planting seems to reduce BSR disease incidence substantially. It is reasonable to assume that microbial competition and antagonism can diminish inoculum loads during this fallow period.
In larger oil palm plantations, infected oil palm trees detected in existing plantation are pushed over, using a backhoe. The stumps and root masses are excavated by digging a hole (2 by 2 by 1 m). The large hole also minimizes the risk of transmitting BSR to neighboring palm trees. The stumps, trunks, and root masses are shredded into small pieces and composted (Chung 2011; Idris et al. 2004b). In North Sumatra, Indonesia, similar methods were applied and BSR incidence 9 years later was only 6.3% (Priwiratama et al. 2020).
In existing oil palm plantations, digging trenches around diseased palm trees has been used to restrict the disease from spreading by root contact. The trenches must be deep enough to prevent the roots from passing beneath them. A trench of 2 by 2 m was effective in slowing down the spread of BSR for up to 14 years (Chung 2011; Hasan and Turner 1998). However, the cost of digging and maintaining trenches makes this practice too expensive to be practical (Turner 1981).
Soil mounding piles soil around the base of the stem up to a height of 75 cm. This method stimulates growth of new roots and helps to prevent trees from being toppled over by wind. Soil mounding in combination with chemical fumigants and organic and inorganic fertilizers lengthened the economic life of infected palm trees by 2 to 3 years (Ho and Hashim 1997; Priwiratama et al. 2014). Removing the outer diseased tissue, followed by soil mounding, can also lengthen the trees’ economic life (Lim et al. 1993). The lesion is removed by chisels; exposed tissues are than treated with protectant chemicals such as coal tar or a mixture of coal tar and thiram. Removing symptomatic trunk tissue can be effective on palm trees over 12 years old but only in the short term; if the lesion is not completely removed, infection may reoccur (Priwiratama et al. 2014; Singh 1990; Turner 1981).
Biological control
Significant efforts have been made in Malaysia and Indonesia to control BSR pathogens, using bacteria and fungi as biocontrol agents. Trichoderma spp. have shown some success in inhibiting the growth of G. boninense in vitro, in plant houses, and under field conditions (Ilias 2000; Nur Ain Izzati and Abdullah 2008; Sariah et al. 2005). Some of the Trichoderma spp. tested were Trichoderma harzianum (Nur Ain Izzati and Abdullah 2008), T. reesei, and T. asperellum (Haryadi et al. 2021). However, Trichoderma is effective against the pathogen only at very early stages of infection (Abdullah et al. 2003).
Other potential fungal biocontrol agents, tested singly or in combination in vitro as well as in nursery trials, have diminished the severity of BSR. These potential biocontrol agents include Scytalidium parasiticum (Goh et al. 2016), a combination of Talaromyces apiculatus and Clonostachys rosea (Goh et al. 2020), and a combination of the mycorrhizal fungus Glomus intraradices with Pseudomonas aeruginosa and G. clarum with P. aeruginosa (Sundram et al. 2015).
Bacterial biocontrol agents such as Burkholderia and Streptomyces spp. suppressed growth of Ganoderma boninense in nursery trials (Maizatul et al. 2012; Shariffah Muzaimah et al. 2012). In an in vitro study, a P. aeruginosa isolate recovered from a soil sample inhibited growth of G. boninense (Lim et al. 2019).
Although several biocontrol agents have shown promising results in nursery assessments, field trials are necessary to determine their effectiveness in field conditions. However, field trials for BSR are time consuming, and a minimum of 3 to 5 years are required to assess effectiveness (Susanto et al. 2005). To overcome this drawback, Flood et al. (2000) developed a bait seedling method wherein seedlings were planted in close proximity to varying concentrations of G. boninense inoculum. This method facilitates conducting field trial observations over a shorter (20 to 24 months) time frame.
Two products containing biocontrol agents that are effective against G. boninense are commercially available in Malaysia. One of the products, known as Hendersonia GanoEF, containing the endophytic fungus Hendersonia toruloidea, was formulated in organic and inorganic fertilizers as a carrier. The formulation with the endophytic fungus can prevent pathogen infection and increase soil fertility and vegetative growth (Idris et al. 2012). Another product, EMBIO actinoPLUS, was developed from the soil actinomycete Streptomyces nigrogriseolus GanoSA1, with a dry powder formulation comprising vermiculite and biochar (Shariffah Muzaimah et al. 2012, 2021). Similarly to Hendersonia GanoEF, EMBIO actinoPLUS also inhibited the growth of G. boninense and reduced infection.
Fertilizer
Mineral nutrition plays a major role in plant disease resistance against pathogens through the production of secondary metabolites, including saponins, glucosinolates, phytoalexin, and terpinoids (Piasecka et al. 2015), and lignification (Engelhard 1989). The severity of plant diseases can be reduced by providing adequate mineral nutrients through proper fertilizer programs. Mineral nutrient supplementation is also one of the strategies to control or manage G. boninense infection. Studies have been performed on infected oil palm trees, and treatment with nutrient fertilizer reduced disease severity and inhibited the growth of BSR pathogens (Bivi et al. 2016; Najihah et al. 2015; Rebitanim et al. 2020; Tengoua et al. 2014).
Foliar application of salicylic acid, copper, and calcium reduced BSR severity and suppressed the growth of G. boninense in infected oil palm seedlings (Bivi et al. 2016). Supplementation with mineral nutrients may restrict pathogen penetration into the host tissues through lignification of the cell wall, particularly thickening of the secondary cell walls and middle lamella (Bivi et al. 2016).
A new fertilizer, GanoCare, was formulated to improve the physiology and growth of oil palm, as well as to prevent BSR infection. The product contains powdered empty oil palm fruit bunches and several minerals, including N, P, K, Ca, Mg, Fe, Si, B, Cu, Zn, and Mn (Rebitanim et al. 2020). Application of GanoCare to oil palm seedlings in nursery and field assessments improved vegetative growth, including height, bulb diameter, and leaf area. Chlorophyll content and the photosynthesis rate also increased. Pretreatment and continuous treatment with GanoCare reduced BSR severity (Rebitanim et al. 2020).
Treatment with Cu, B, and Mn combined with N-P-K-Mg fertilizer reduced BSR severity in oil palm seedlings (Tengoua et al. 2014). The combination of two micronutrients (B+Cu, B+Mn, and Cu+Mn) also reduced incidence and severity of BSR in oil palm seedlings (Tengoua et al. 2014). However, field studies have not been performed to determine the efficacy of micronutrient treatment in oil palm trees.
Five forms of silicon (calcium silicate, potassium silicate, sodium silicate, silicon oxide, and sodium meta-silicate) at four concentrations (0, 800, 1,200, and 2,000 mg liter−1) were used as supplemental fertilizers for oil palm seedlings. After 8 months, the seedlings supplemented with silicon oxide at 1,200 mg liter−1 showed 53% disease severity, compared with 95% disease severity for the nontreated control (Najihah et al. 2015). As assessed from the foliar symptoms and infection in the roots and bole, silicon fertilization restricted G. boninense colonization of the host tissues and, thus, reduced the extent of root infection and development of lesions in the stem.
Resistant varieties
Resistant or tolerant oil palm trees may provide a long-term control method against BSR. However, the development of resistant or tolerant varieties is hindered by inadequate plant genetic resistance sources and absence of rapid and effective screening methods (Soh 2017). Moreover, oil palm has a narrow genetic base, with available genetic resources only from E. guineensis originating from Africa and E. oleifera from South America. These species are crossable and used in genetic improvement programs as a source of diversity (Le Meunier 1975; Rajanaidu 1986). Regarding the development of resistant or tolerant oil palm planting materials, differences in susceptibility to BSR were detected within both E. guineensis and E. oleifera (de Franqueville et al. 2001; Durand-Gasselin et al. 2005), which paved the way for using both species as genetic resources.
To distinguish the sources of susceptibility and resistance, early screening tests are conducted at the prenursery stage or at the nursery using artificial inoculation of the pathogen (Breton et al. 2006, 2009). Artificial inoculation includes germinated seed inoculation (Breton et al. 2006) and root inoculation (Ariffin and Idris 1991). Several parameters used in the screening test have been standardized, including the oil palm planting material, physiological stage, shade during the prenursery trial, temperature, inoculum source size, and incubation period (Breton et al. 2006). The duration of the early screening tests should be as short as possible to evaluate a large number of oil palm progenies or planting materials.
Partial resistance of oil palm to BSR has been reported from the early screening of planting materials in Malaysia and Indonesia. Idris et al. (2004a) tested 12-month-old seedlings from 23 oil palm progenies, and partially resistant progenies were found in the crosses of Congo and Cameroon germplasm sources. Partial resistance was evident from reduced severity of foliar symptoms and slow progression of root and stem infection. Partial resistance was also reported by Durand-Gasselin et al. (2005) and Breton et al. (2006) in their studies on private oil palm plantations in Indonesia. The source of resistance reported by Durand-Gasselin et al. (2005) was the Mangenot line of E. oleifera. The results of these studies are very promising and can lead to the development of Ganoderma-resistant varieties.
Resistant oil palm development is hampered by limited information on defense-related genes. In a study by Tisné et al. (2017), four Ganoderma resistance loci were identified based on the mapping of multiparental oil palm populations from nine major founders of ongoing breeding programs in Indonesia and Benin. Two of the identified resistance loci affected occurrence of BSR symptoms and two loci were associated with the rate of death of the oil palm tree. The study was performed using a QTL mapping method and generated important data for the selection of oil palm varieties resistant to BSR. Although the results appear promising, it is unlikely to provide an immediate solution for developing resistant or tolerant planting materials.
Conclusion and Future Outlook
Studies on BSR conducted since the 1960s have improved our understanding of the occurrence, infection, and development of this disease. G. boninense is the main causal pathogen of BSR, particularly in Malaysia, Indonesia, and Papua New Guinea, although other species have also been reported. Biotrophic and necrotrophic phases of the pathogen have been revealed through gene expression evaluation of the infected oil palm and histological studies of infection. Gene expression studies revealed the molecular defense mechanisms of oil palm against BSR, which is useful for understanding the underlying resistance mechanisms of the oil palm. This information can also be used in the screening and development of BSR-resistant or -tolerant oil palm.
To date, various studies have focused on the early detection of BSR in oil palm, particularly in Malaysia and Indonesia. Several detection methods or tools have been developed. However, some of these methods lack information on the effectiveness when applied in the field, because they have been tested only in laboratory or plant house trials. Moreover, many of these methods require specialized knowledge to handle the equipment and analyze the data before they can be applied in decision making on disease management. Nevertheless, development of early detection methods and tools for BSR could aid in improving the management of BSR infections.
In addition to diagnostic tools, several products, including biofertilizers, nutrient supplementation, and products containing biocontrol agents, have also been developed and are commercially available. These products can help reduce the risk of G. boninense infection and avoid potentially higher yield losses.
Currently, BSR is considered manageable in Malaysia and Indonesia through integrated BSR disease management. The main aim of integrated BSR management is to minimize the occurrence of this disease, particularly in replanting plantations, because high disease incidence commonly occurs in these fields. Integrated BSR management is also aimed at extending the productive life of oil palm trees. Until a more effective method to control G. boninense infection is developed, the existing integrated BSR management strategy includes cultural methods such as sanitation, clean clearing, and application of fungicides.
Studies on the development of resistant or tolerant oil palm trees are at an early stage, and there is still a long way to go. Germplasm collections of oil palm from Africa and South America are available in Malaysia, and partial resistance against BSR has been reported to facilitate the breeding of resistant or tolerant oil palm trees. Moreover, the oil palm genome has been sequenced (Singh et al. 2013), which will provide an opportunity to determine genes associated with disease resistance and to identify marker alleles useful for breeding purposes.
Dr. Latiffah Zakaria is a professor of plant pathology at the School of Biological Sciences, Universiti Sains Malaysia (USM), Penang, Malaysia. She received her B.Sc. (Biology) and Ph.D. (Plant Pathology) from Universiti Putra Malaysia. Her academic career is split among teaching, research, and administration. She has been appointed as Agrobiology Programme Chairman, Manager of Scientific Equipment and Services, Deputy Dean of Industry and Community Network and currently is the Deputy Dean of Postgraduate, Innovation and Industry-Community Engagement. Her research areas are on various aspects of plant pathology and mycology, particularly those related to systematics, pathogenicity, and mycotoxigenicity of plant-pathogenic fungi, endophytic fungi, and spoilage fungi.
Jakarta – Jamur Mikoriza merupakan salah satu sumber daya yang dapat membantu meningkatkan kelayakan hidup manusia. Hal ini dikarenakan hubungan jamur dan tumbuhan di dalam tanah dapat membantu mengendalikan emisi karbon dan transportasi karbon.
Peran Jamur Mikoriza dalam Pengendalian Emisi Karbon
Dilansir dari detikEdu yang mengutip dari laman BBC Science Focus, tim peneliti asal Inggris, Afrika Selatan, dan Belanda mengungkapkan bahwa tumbuhan mampu menyerap sekitar 13,12 gigaton karbon dioksida dari atmosfer ke dalam jamur mikoriza setiap tahunnya. Karbon yang diserap ke dalam tanah setara dengan 36% emisi bahan bakar fosil global tahunan.
Penyerapan ini terjadi karena terdapat hubungan simbiotik yang saling menguntungkan antara jamur mikoriza dan akar tanaman. Dalam proses ini, jamur akan menyediakan nutrisi yang penting bagi pertumbuhan tanaman dan akan mengambil sebagian gula serta karbon dioksidanya yang diproduksi saat berfotosintesis.
“Penelitian ini memberi kita kesadaran akan peran potensial yang dimainkan oleh jamur dalam mengendalikan CO2 yang meningkat,” jelas seorang peneliti interaksi tanah-tanaman-mikroba di University of Cape Town, Afrika Selatan, Dr Heidi Hawkins.
Ia juga menambahkan bahwa saat ini fokus utama ditempatkan pada perlindungan dan restorasi hutan untuk mengurangi perubahan iklim. Akan tetapi, jumlah karbon dioksida yang dipindahkan dari atmosfer selama proses fotosintesis yang dikirim ke bawah tanah menuju jamur mikoriza hanya sedikit.
Peran Jamur dalam Transportasi Karbon
Meskipun tumbuhan dan jamur mikoriza memiliki hubungan simbiotik yang saling menguntungkan, baru-baru ini para ilmuwan juga menemukan bahwa jamur juga bertindak sebagai pintu gerbang agar karbon mampu masuk ke dalam tanah.
Jamur mikoriza akan mengangkat nutrisi dan mineral melalui tanah menggunakan jaringan berfilamen bernama miselium. Lalu, jaringan miselium akan diperluas menggunakan karbon yang mereka dapatkan.
Diketahui, hal tersebut mampu memperkuat struktur tanah. Sayangnya, hingga kini para peneliti belum mengetahui berapa lama karbon bisa tinggal di dalam jamur.
“Walaupun jamur mikoriza tentu saja mengandung dan melepaskan karbon ke dalam tanah, saat ini kami belum tahu apakah jamur mikoriza adalah simpanan karbon (mempertahankan karbon) atau penyerap karbon (meningkatkan karbon dari waktu ke waktu),” jelas Hawkins.
“Namun demikian, sudah ada beberapa tindakan praktis yang dapat kita lakukan, misalnya dengan melestarikan daerah-daerah di mana kita tahu bahwa asosiasi mikoriza tertentu tersebut menghasilkan alokasi karbon yang tinggi ke dalam tanah (hutan dan heathland),” tambahnya.
Selanjutnya, para peneliti juga menekankan tentang perlunya dilakukan penelitian lebih lanjut. Hal ini dikarenakan Organisasi Pangan dan Pertanian Perserikatan Bangsa-Bangsa atau UN Food and Agriculture Organisation (FAO) telah memperingatkan bahwa 90% tanah dapat terdegradasi pada tahun 2050 mendatang.
“Banyak kegiatan manusia yang merusak ekosistem di bawah tanah. Selain membatasi kerusakan, kita perlu meningkatkan tingkat penelitian secara radikal,” pungkas penulis buku ‘Entangled Life: How Fungi Make Our Worlds, Change Our Minds And Shape Our Futures’ Merlin Sheldrake.
Diakui atau tidak tanaman kelapa sawit merupakan tanaman yang berharga. Lantaran berkaitan dengan tingginya keuntungan ekonomi dari penjualan komoditas tersebut, dimana Indonesia merupakan produsen terbesar di dunia dimana Pulau Sumatera merupakan saah satu wilayah terpenting untuk produksi minyak sawit.
Dalam sebuah penulisan ilmiah berjudul, “Ganoderma boninense Disease of Oil Palm to Significantly Reduce Production After 2050 in Sumatra if Projected Climate Change Occurs”, yang diterbitkan pada 2019 lalu mencatat, pulau Sumatera dapat dianggap sebagai model bagi wilayah perkebunan kelapa sawit lainnya di Asia Tenggara.
Dimana, luas wilayah di Sumatera yang memiliki iklim yang cocok untuk menanam kelapa sawit akan berkurang akibat proyeksi perubahan iklim seperti yang ditunjukkan secara khusus dalam riset yang dilakukan R Russel M Paterson, periset Universitas Minho, Portugal.
Catat Paterson, iklim yang semakin tidak sesuai akan menyebabkan peningkatan penyakit busuk pangkal batang (BSR) yang muncul karena infeksi jamur Ganoderma boninense, seperti yang diperkirakan sebelumnya, yang menjadi perhatian utama terhadap keberlanjutan di Asia Tenggara.
Dalam sebuah pendekatan baru dijelaskan bahwa dimana (a) dilakukan penentuan iklim yang sesuai untuk pertumbuhan kelapa sawit di Sumatera dan (b) pengurangan untuk menentukan tingkat BSR di masa depan di pulau tersebut.
Nyatanya, ketidaksesuaian iklim untuk kelapa sawit diperkirakan akan meningkatkan serangan jamur ganoderma secara drastis setelah tahun 2050, yang mana serangannya mencapai sangat tinggi di sebagian besar wilayah pulau tersebut.
Dilansir InfoSAWITdari PubMed Central (PMC), bahkan kemungkinan besar akan membuat produksi minyak sawit menjadi tidak berkelanjutan pada tahap tertentu antara tahun 2050 dan 2100.
“Sumatera Utara mungkin lebih berkelanjutan dibandingkan wilayah lain di Sumatera. Dampak perubahan iklim yang diperkirakan memerlukan perbaikan sebelum tingginya tingkat serangan ganoderma dan iklim yang tidak cocok untuk kelapa sawit bisa terwujud,” demikian catat riset tersebut.