BBC Science Focus, tim peneliti asal Inggris, Afrika Selatan, dan Belanda mengungkapkan bahwa tumbuhan mampu menyerap sekitar 13,12 gigaton karbon dioksida dari atmosfer ke dalam jamur mikoriza setiap tahunnya. Karbon yang diserap ke dalam tanah setara dengan 36% emisi bahan bakar fosil global tahunan. Dunia Sawit pasti sudah mengenal siapa sih Mikoriza itu? Apalagi si jamur ini heboh setelah beberapa lahan sawit terkena serangan Ganoderma. Tak kenal maka tak sayang, yuk kita mengenal lebih jauh tentang Mikoriza ini. Nama “mikoriza” adalah serapan dari istilah bahasa Inggris, mycorrhiza, yang juga bentukan dari dua kata bahasa Yunani Kuno: μύκης mýkēs, “jamur”, dan ῥίζα rhiza.
Selain bermanfaat sebagai mikoriza penyubur tanaman, jamur kapang juga digunakan sebagai ragi pengolah makanan seperti pembuatan roti, tempe, oncom, minuman anggur brewery, dan tapai serta pengobatan antibiotik. sedangkan Kelompok yang menyebabkan keracunan jamur adalah mikotoksin.
Mikoriza adalah cendawan (fungi) yang bersimbiosis dengan tumbuhan. Biasanya simbiosis ini terletak di sistem perakaran tumbuhan. Ada juga cendawan yang bersimbiosis dengan cendawan lainnya, tetapi sebutan mikoriza biasanya ditunjukan untuk cendawan yang melakukan simbiosis dengan tumbuhan. Bentuk simbiosis ini terutama adalah simbiosis mutualisme, meskipun pada beberapa kasus dapat berupa simbiosis parasitisme lemah.
Mikoriza memerlukan akar tumbuhan untuk melengkapi daur hidupnya. Sebaliknya, beberapa tumbuhan bahkan ada yang tergantung pertumbuhannya dengan mikoriza. Beberapa jenis tumbuhan tidak tumbuh atau terhambat pertumbuhannya tanpa kehadiran mikoriza di akarnya. Sebagai misalnya, semaian pinus biasanya gagal tumbuh setelah pemindahan apabila tidak terbentuk jaringan mikoriza di sekitar akarnya.
Mikoriza dapat diinokulasi secara buatan. Namun, inokulasi mikoriza asing memerlukan bantuan mikoriza lokal, misalnya dengan menambahkan tanah dari tempat asal tumbuhan.
Dua kelompok mikoriza terbesar adalah ektomikoriza (EcM) dan endomikoriza (EM). Endomikoriza terutama didominasi oleh mikoriza arbuskular (arbuscular mycorrhizae, AM), ditambah dengan sekelompok mikoriza erikoid dan mikoriza arbutoid yang menginfeksi tumbuhan kelompok Ericoidae.
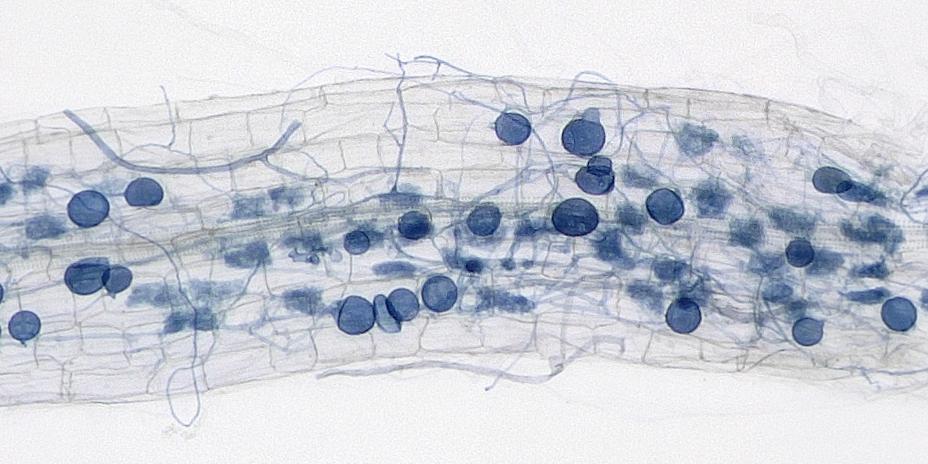
Semua endomikoriza termasuk ke dalam filum Glomeromycota, misalnya genus Gigaspora, Scultellospora, Acaulospora, Entrophospora, Glomus, dan Sclerocystis. Terdapat sekitar 150 jenis (spesies) spora cendawan AM yang telah dideskripsi. AM tergolong dalam kelompok khusus dari populasi mikoriza yang sangat banyak mengkolonisasi rizosfer, yaitu di dalam akar, permukaan akar, dan di daerah sekitar akar. Hifa eksternal yang berhubungan dengan tanah dan struktur infeksi seperti arbuskula di dalam akar menjamin adanya perluasan penyerapan unsur-unsur hara dari tanah dan peningkatan transfer hara (khususnya fosfor) ke tumbuhan, sedangkan cendawan memperoleh karbon organik dari tumbuhan inangnya (Marschner, 1995).
Mikoriza dibagi menjadi dua kelompok berdasarkan cara menginfeksinya, yaitu ektomikoriza dan endomikoriza. Ektomikoriza menginfeksi permukaan luar tanaman dan di antara sel-sel ujung akar. Akibat serangannya, terlihat jalinan miselia berwarna putih pada bagian rambut-rambut akar, dikenal sebagai jala Hartig. Serangan ini dapat menyebabkan perubahan morfologi akar. Akar-akar memendek, membengkak, bercabang dikotom, dan dapat membentuk pigmen. Infektivitas tergantung isolat dan kultivar tumbuhan inang. Tumbuhan inangnya biasanya tumbuhan tahunan atau pohon. Beberapa di antaranya merupakan komoditas kehutanan dan pertanian seperti sengon, jati, serta beberapa tanaman buah seperti mangga, rambutan, dan jeruk. Selain itu pohon-pohon anggota Betulaceae, Fagaceae, dan Pinaceae juga menjadi inangnya. Pada umumnya ektomikoriza termasuk dalam filum Basidiomycota dan Ascomycota. Ada sedikit anggota Zygomycota yang juga menjadi cendawan ektomikoriza.
Endomikoriza menginfeksi bagian dalam akar, di dalam dan di antara sel-sel ujung akar. Hifa masuk ke dalam sel atau mengisi ruang-ruang antarsel. Jenis mikoriza ini banyak ditemukan pada tumbuhan semusim yang merupakan komoditas pertanian penting, seperti kacang-kacangan, padi, jagung, beberapa jenis sayuran dan tanaman hias. Infeksi ini tidak menyebabkan perubahan morfologi akar, tetapi mengubah penampilan sel dan jaringan akar. Berdasarkan tipe infeksinya, dikenal tiga kelompok endomikoriza: ericaceous (Ericales dengan sejumlah Ascomycota), orchidaceous (Orchidaceae dengan sekelompok Basidiomycota), dan vesikular arbuskular (sejumlah tumbuhan berpembuluh dengan Endogonales, membentuk struktur vesikula (gelembung) dan arbuskula dalam korteks akar) disingkat MVA.
Mikoriza arbuskular (AM, dulu disebut mikoriza vesikular-arbuskular, VAM) tumbuh dari luar perakaran lalu masuk ke dalam jaringan perakaran dan pada gilirannya memasuki sel-sel perakaran. AM di dalam jaringan akan membentuk arbuskula, yaitu jaringan hifa yang menembus sela-sela sel dan bahkan menembus sel melalui plasmalema. Di dalam sel, hifa akan membentuk vesikula, suatu gelembung-gelembung kecil di sitoplasma. AM sulit ditumbuhkan secara aksenik (media buatan) sehingga MVA dianggap merupakan simbion obligat (wajib).
Vesikula berbentuk butiran-butiran di dalam sitoplasma yang mengandung lipid dan menjadi alat reproduksi vegetatif mikoriza, khususnya bila sel pecah akibat rusaknya korteks akar. Arbuskula berwujud kumpulan hifa yang menembus plasmalema dan membantu transportasi hara di dalam sel tumbuhan. Pembentukan vesikula dan arbuskula dalam sel menunjukkan bahwa simbiosis telah terjadi dengan sempurna dan tanaman sudah dapat menikmati hasil kerja sama dengan mikoriza berupa meningkatnya ketersediaan unsur hara yang diserap dari dalam tanah.
Selain vesikula dan arbuskula, terbentuk hifa eksternal yang dapat membantu memperluas ruang penyerapan hara oleh akar. Pada bawang merah, misalnya, panjang hifa eksternal dapat mencapai 80 cm per satu cm panjang akar. Di luar akar, hifa dapat membentuk sporangium yang menghasilkan spora sebagai alat reproduksi.
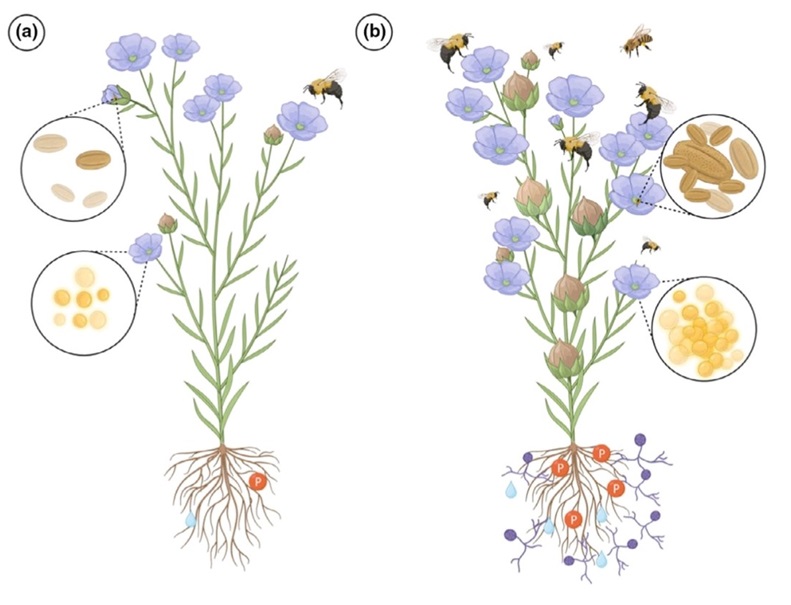
AM banyak membawa keuntungan bagi tumbuhan simbionnya. Ia memperbaiki hasil tumbuhan dan mengurangi masukan pupuk pada tanaman pertanian. Ini terjadi karena MVA meningkatkan ketersediaan beberapa hara di tanah yang diperlukan tanaman, terutama fosfat. Peningkatan penyerapan fosfat diiringi dengan peningkatan penyerapan hara lain, seperti nitrogen (N), seng (Zn), tembaga (Cu), dan belerang (S).
Selain itu, MVA memperluas ruang tanah yang dapat dijangkau oleh tanaman inang. Jeruk, umpamanya, dikenal responsif terhadap inokulasi MVA. Inokulasi ini dapat mengarah pada menurunnya penggunaan pupuk P. Selain meningkatkan ketersediaan hara, AM meningkatkan toleransi tumbuhan terhadap kurangnya pasokan air. Luasnya jaringan hifa di tanah membantu akar menyerap air. MVA memengaruhi ketahanan tumbuhan inang terhadap serangan penyakit. AM, tergantung jenisnya, dapat mengurangi pengaruh serangan jamur patogen. Demikian pula, juga dapat mengurangi serangan nematoda. Sebaliknya, tumbuhan yang terinfeksi AM menurun ketahanannya terhadap serangan virus.
Pengaruh AM lain yang pernah teramati adalah dukungannya terhadap simbiosis antara bakteri bintil akar dan polong-polongan, produksi giberelin oleh Gibberella mosseae, memengaruhi sintesis fitohormon tertentu, dan memperbaiki struktur agregasi tanah.
Manfaat Umum MVA
Manfaat dari MVA dapat dikelompokkan menjadi tiga, yaitu manfaat dalam ekosistem, manfaat bagi tanaman, dan manfaatnya bagi manusia. Manfaat mikoriza MVA dalam ekosistem sangat penting, yaitu berperan dalam siklus hara, memperbaiki struktur tanah dan menyalurkan karbohidrat dari akar tanaman ke organisme tanah yang lain, sedangkan manfaat bagi tanaman yaitu dapat meningkatkan penyerapan unsur hara, terutama P. MVA ini dapat mengeluarkan enzim fosfatase dan asam-asam organik, khususnya oksalat yang dapat membantu membebaskan P. MVA dapat membantu mengatasi masalah ketersediaan fosfat melalui dua cara, pengaruh langsung melalui jalinan hifa eksternal yang diproduksinya secara intensif sehingga tanaman bermikoriza akan mampu meningkatkan kapasitasnya dalam menyerap unsur hara dan air dan pengaruh tidak langsung, dimana mikoriza dapat memodifikasi fisiologis akar sehingga dapat mengeksresikan asam-asam organik dan fosfatase asam ke dalam tanah. Fosfatase asam merupakan suatu enzim yang dapat mamacu proises mineralisasi P Organik dengan mengkatalisis pelepasan P dari kompleks organik menjadi kompleks anorganik. Manfaat lain yaitu bagi manusia, mikoriza dapat meningkatkan produktivitas tumbuhan, karena dapat memproduksi bunga lebih awal.
Mekanisme Penyerapan Fosfat oleh Mikoriza
Peranan MVA tersebut dalam meningkatkan ketersediaan dan serapan P dan unsur hara lainnya melalui proses sebagai berikut:
- Modifikasi Kimia oleh mikoriza dalam proses kelarutan P tanah Pengaruh Mikoriza Arbuskula Pada Ketersediaan dan Penyerapan Unsur Hara Pada tahap ini, terjadi modifikasi kimia oleh mikoriza terhadap akar tanaman, sehingga tanaman mengeksudasi asam-asam norganik dan enzim fosfatase asam yang memacu proses mineralisasi P. Eksudasi akar tersebut terjadi sebagai respon tanaman terhadap kondisi tanah yang kahat P, yang mempengaruhi kimia rizosfer.
- Perpendekan jarak difusi oleh tanaman bermikoriza. Mekanisme utama bagi pergerakan P ke permukaan akarah melalui difusi yang terjadi akibat adanya gradien konsentrasi, serta merupakan proses yang sangat lambat. Jarak difusi ion-ion fosfat tersebut dapat diperpendek dengan hifa eksternal CMA, yang juga dapat berfungsi sebagai alat penyerap dan translokasi fosfat.
- Penyerapan P tetap terjadi pada tanaman bermikoriza meskipun terjadi penurunan konsentrasi minimum P. Konsentrasi P yang ada di larutan tanah dapat menjadi sangat rendah dan mencapai konsentrasi minimum yang dapat diserap akar, hal ini terjadi sebagai akibat terjadinya proses penyerapan ion fosfat yang ada di permukaan akar. Di bawah konsentrasi minimum tersebut akar tidak mampu lagi menyerap P dan unsur hara lainnya, sedangkan pada akar bermikoriza, penyerapan tetap terjadi sekalipun konsentrasi ion fosfat berada di bawah konsentrasi minimum yang dapat diserap oleh akar. Proses ini ini terjadi karena afinitas hifa eksternal yang lebih tinggi atau peningkatan daya tarikmenarik ion-ion fosfat yang menyebabkan pergerakan P lebih cepat ke dalam hifa MVA.
Dadang Gusyana
Ilmuwan – Regional Agronomist